|
 Летеће рибе (породица Екоцоетидае), свима добро познате из описа поморских путовања, саставни су део пејзажа топлих мора и једна су од његових најкарактеристичнијих спољних манифестација. Летеће рибе (породица Екоцоетидае), свима добро познате из описа поморских путовања, саставни су део пејзажа топлих мора и једна су од његових најкарактеристичнијих спољних манифестација.
У екосистему тропских вода отвореног океана летеће рибе заузимају јединствени положај, будући да су једине масивне планктофаге које непрестано насељавају површинске слојеве епипелагијске зоне (горњи слој воденог стуба).
Летеће рибе саме по себи чине важну компоненту исхране предаторских риба - корифена, скуша, мале туне, као и морских птица, лигње и делфини. У неким областима (Јапан, Филипини, Индија, Полинезија, острва Кариба) спроводи се посебан риболов летеће рибе, који, међутим, има само локални значај, али даје, према грубој процени, најмање 500 хиљаду центара годишње. Летећа риба се лови шкржним мрежама, ташном и фиксним мрежама, мрежама за капице и посебним замкама и рибарским штаповима; постоје и друге методе засноване на посебностима екологије ових риба (посебно на њиховој позитивној реакцији на вештачко светло и на прилазима мријеста обалама).
Летеће рибе, како им само име говори, могу да лете ваздухом. Одакле долази ова способност? Сви представници реда гарфисх, који укључују летеће рибе, а поред њих, такође полу-рибе, гарфисх и саури, настањују се у најгорњим слојевима воде. Многи од њих, уплашени или у потрази за пленом, могу искочити из воде, понекад правећи читав низ узастопних скокова, попут камена за рикошетирање. Побољшање ових скокова на крају је довело до клизног лета, омогућавајући летећим рибама да побегну од многих грабежљиваца, мада им, наравно, не гарантује потпуну сигурност: на пример, корифен, уплашивши летећу рибу, гони је под водом и хвата је у тренутку кад тоне у воду. Ипак, објашњење лета као уређаја за спасавање од предатора сада је опште прихваћено и већ дуго се не доводи у питање. Друго становиште износи проф. В.Д.Лебедев, који сматра да су миграције крме уз употребу сталних ветрова играле одлучујућу улогу у развоју лета. Међутим, мора се рећи да још увек није доказано постојање миграција летећих риба на велике даљине у самом тропском појасу. Доступни подаци, напротив, сведоче о „седећем“ начину живота.
Летеће рибе су веома разноврсне - породица укључује 7 родова и око 60 врста. Способност летења изражена је у различитом степену код различитих родова. Лет „примитивних“ летећих риба из родова Фодиафор и Парекоцоетус, са релативно кратким прсним перајама, мање је савршен него код риба са дугим „крилима“. Еволуција лета летеће рибе одвијала се, очигледно, у два смера. Један од њих је довео до формирања рода Екоцоетус - „двокрилне“ летеће рибе које у лету користе само прсне пераје које у њима достижу врло велике величине (до 80% дужине тела). Други правац представљају „четверокриле“ летеће рибе (4 рода подфамилије Ципселуринае — Прогхицхтхис, Ципселурус, Цхеилопогон, Хирундицхтхис, - комбинујући око 50 врста). Лет ове рибе се изводи помоћу два пара носећих равни: повећале су не само прсне, већ и карличне пераје, а у фази развоја младица обе имају приближно исту површину. Оба смера у еволуцији лета довела су до формирања специјализованих облика добро прилагођених животу у епипелагији.Поред развоја „крила“, прилагођавање лету одразило се и на летеће рибе у структури репне пераје чији су зраци круто повезани једни с другима, а доња оштрица је веома велика у поређењу са горњом , у необичном развоју бешике за пливање, која се наставља испод кичме до самог репа, и у бројним другим карактеристикама. Лет летеће рибе „четворокрилац“ достиже највећи домет и трајање. Развивши значајну брзину у води (око 30 км / х), таква риба искаче на површину мора и неко време, понекад не дуго, клизи по њој раширених прсних пераја-крила, снажно убрзавајући кретање вибрацијама доње оштрице репне пераје уроњене у воду и повећавајући брзину на 60-65 км / х. Тада се риба одваја од воде и, отварајући карличне пераје, планира преко своје површине. У неким случајевима, док лети, летећа риба понекад репом додирује воду и вибрирајући њиме добија додатну брзину. Број таквих додира може досећи три до четири, а у овом случају се трајање лета, природно, повећава. Обично је летећа риба у ваздуху не више од 10 секунди. и лети за то време неколико десетина метара, али понекад се трајање лета повећава на 30 секунди, а његов домет достиже 200, па и 400 м. Очигледно, трајање лета зависи од атмосферских услова, јер у присуству слабог ветра или узлазног ваздушне струје ... летеће рибе лете на велике даљине и дуже остају у лету.
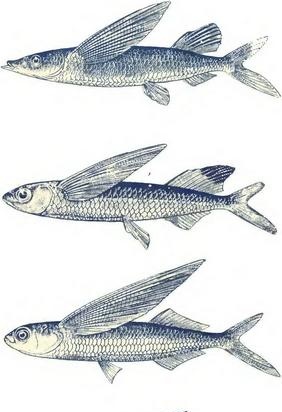
Летеће рибе (од врха до дна): Фодиатор ацутус, Парекоцоетус брацхип-терус, Екоцоетус волитанс (диптеран летећа риба) и Х. спецулигер (четверокрила гута риба).
Многи морнари и путници, посматрајући летећу рибу са палубе брода, тврдили су да су „јасно видели да риба маше крилима баш попут вретенца или птице“. У ствари, „крила“ летећих риба остају готово непомична током лета и не врше никакво клацкање. Само се угао нагиба пераја може, очигледно, произвољно променити, а то омогућава риби да мало промени смер лета. Дрхтање пераја, које је посматрачу приметно, највероватније је само последица лета, али нимало његов узрок. Објашњава се једноставном вибрацијом раширених „крила“, која је посебно јака у оним тренуцима када риба, већ у ваздуху, наставља да ради у води репном перајом.
Током проучавања летећих риба Атлантског океана, познати дански ихтиолог Антон Брун (АФ Бруун. Летеће рибе (Еко-цоетидае) Атлантика, „Дана Репорт“, 1935, бр. 6.) прво је приметио да је ова група која се увек сматрао карактеристичним за отворени океан, садржи не само океанске, већ и неритске (приморске) облике. Брун је такође приметио да породица укључује тропске ("екваторијалне", у његовој терминологији) врсте које се не налазе изван самог тропског појаса и суптропске врсте које живе само на ивицама ове зоне. Према његовом мишљењу, температура површинских слојева воде је фактор који ограничава ширење летеће рибе. Даље проучавање екологије ове групе показало је да подела летећих риба на океанске и неритске групе донекле поједностављује стварно стање. Поред чисто неритских врста и врста ограничених на отворене воде, постоји и псеудо-океанска, односно неритско-океанска, врста која се јавља далеко од обале само у неком периоду свог животног циклуса.
Подела летеће рибе у ове групе одређена је еколошким разликама. Неритске врсте се обично размножавају полагањем прилепљених јаја на тврду подлогу (алге, дно). Типични представници ове групе укључују Фодиатор ацутус, Парекоцоетус менто, неки представници рода Ципселурус и низ других врста. Насупрот томе, океанске летеће рибе (све врсте рода Екоцоетуснеки Цхеилопогон, Прогницхфхис и Хирундицхтхис) насељавају само отворена подручја, а њихова јаја се или развијају у воденом ступцу, или се таложе на плутајућим објектима који се увек могу наћи у мору (заносеће алге, пераја, птичје перје). Коначно, псеудо-океанске врсте (ту спада и већина врста које углавном припадају роду Сурселурус и Цхеилопогон) могу постојати у отвореном океану, али им је за размножавање потребан чврст приобални супстрат. Станишта неритских и океанских летећих риба значајно се разликују у равнотежи сезонских трофичних циклуса заједница које их насељавају. Чињеница је да је у отвореним водама тропског океана производња фитопланктона дуго времена близу потрошње зоопланктона, а производња наредних нивоа близу је предаторима на вишим нивоима система исхране. Стога су океанске пелагичне заједнице међу најуравнотеженијим у погледу трофичних циклуса и просторне хомогености дистрибуције организама. За разлику од ових заједница, у неритским регионима производња дуго премашује пашу, а биоценозе које их насељавају нису уравнотежене у смислу трофизма. Пелагичне животиње су овде дистрибуиране врло неравномерно због „пегавог“ цветања алги и чине школе и школе.
Све летеће рибе су стенотермалне, односно живе у прилично уском температурном опсегу, константном за сваку врсту. Они су мање или више термофилни, а већина врста се не јавља или се готово не јавља на температури воде нижој од 23 °. Ове врсте чине тропску групу. Само су се многи чланови породице прилагодили животу у суптропским водама на температурама од 18–20 ° и ниже, а лети продиру чак и у умерене регионе; минимална температура на којој су сретене „најотпорније“ врсте - Хирундицхфхис ронделетии, је само 15,5 °. Субтропска група укључује само 6-7 врста летећих риба (тј. Само око 10% свих врста породице). У суптропским водама налазе се само високоспецијализоване летеће рибе, док представници примитивних родова Фодиафор и Регехосоетус живе само у тропском појасу.
Географска дистрибуција неритских и неритно-океанских летећих риба у потпуности подлеже свим законима који регулишу дистрибуцију тропских приобалних риба уопште. Препрека њиховом насељавању нису само континенталне баријере, већ и отворене воде, посебно „фаунистичка баријера источног Пацифика“ - регион без острва Тихог океана, између обала Америке и крајњих источних архипелага Полинезије. Управо овај разлог објашњава значајну разлику у фаунима летећих риба у западним и источним деловима Тихог океана. Распрострањеност неритних врста, по правилу, је релативно мала због разноликости еколошких услова у близини обале, а међу њима често постоје врло уски ендеми који живе на врло ограниченим подручјима. Фактори који ограничавају дистрибуцију ове рибе дуж обале су температура воде, њен салинитет (готово све врсте избегавају освежена подручја), капацитет исхране региона и, вероватно, такође природа дна и присуство вегетације у приморски појас. Примери ове врсте су прилично бројни: постоје врсте ендемске за воде Јужног Јапана и Кореје, за воде Индонезије и суседних региона, за пацифичке воде Централне Америке итд. Суптропска врста, џиновска летећа риба, је врло занимљиво. Цхеилопогон пиннати барбатус, дужине до 50 цм, настањује обалне воде Јапана, Калифорније, северозападне Африке и Шпаније на северној хемисфери и у водама Чилеа, Новог Зеланда, Јужне Аустралије и Јужне Африке на југу. Распон ове врсте показује изузетну сличност са подручјем распрострањења сардина из родова Сардина и Сардинопс... Прекинута дистрибуција у тропском појасу такође је карактеристична за такве летеће рибе као што су Цх. хетерурус и Цх. агоо... Концепт биполарности прилично је применљив на распрострањеност свих ових врста (биполарност овде значи дистрибуцију животиња у умереним или суптропским водама северне и јужне хемисфере у одсуству истих у самој тропској зони) ако се сматра нешто шири смисао од ЛС Берга, који је у своје време приметио да су организми умерених географских ширина биполарни. У данашње време постоји много примера „биполарне“ (или, у терминологији америчких аутора, „антитропске“) дистрибуције у суптропским животињама.
Океанске летеће рибе, по правилу, имају врло проширене, често чак и окологлобалне домете, а њихов распоред највероватније одређује само једна температура површинског слоја воде. Неке врсте имају врло уски распон оптималних температура и зато се налазе само у најтоплијим водама или, напротив, у мање загрејаним водама тропског појаса. Ове врсте укључују, на пример, тихоокеанску популацију врсте Е. волитанс, који се овде налазе на температури од 22-29 °, али најчешћи на 24-28 °. Као резултат, подручје распрострањења ове рибе у најтоплијем западном делу Тихог океана прекинуто је у готово екваторијалној зони на око 15 ° географске ширине и у централним и источним деловима океана, где су екватору температура у површинском слоју је спуштена услед пораста дубоких вода, такав прекид није. Насељавајући јужну периферију саме тропске зоне у Тихом океану Е. обфусиросфрис има посебно уске температурне границе дистрибуције у југоисточном делу свог подручја. Као што показују резултати 4. путовања истраживачког брода Академик Курцхатов, ова летећа риба се лови само у уском појасу вода омеђених изотермама од 19 ° и 22-23 °.
Од посебног интереса је дистрибуција једине суптропске врсте међу представницима океанске групе летећих риба - Хирундицхтхис ронделетии, која има биполарно подручје. Ову рибу очигледно карактеришу сезонске миграције: на северозападу Тихог оцеана мрест се јавља зими између 21 ° и 30 ° Н. сх. на температури воде од 18-23 °, у пролеће почиње кретање на север ради това (у овом тренутку се рибе налазе на температури од 15-17 °), а на јесен - обрнута миграција на јужни део опсег.
Квантитативна дистрибуција летећих риба унутар окупираног подручја одређена је првенствено количином доступне хране, односно обиљем зоопланктона у површинском слоју океана. С тим у вези, дистрибуција летећих риба у различитим деловима тропског региона веома је хетерогена. Подручја отвореног океана, која се одликују највећим концентрацијама летеће рибе, налазе се, по правилу, у близини зона разилажења, где се дешава успон на површину дубоких вода богатих биогеним солима и бележи се повећана биолошка продуктивност. У овом случају, највећа концентрација летеће рибе обично се примећује на одређеној удаљености од одступања. Чињеница је да су врхови обиља сваког следећег Чак и у трофичном ланцу (фитопланктони -> биљоједни зоопланктони -> предаторски планктони -> - планктофажне рибе) донекле померени низводно у односу на максималну бројност претходне везе. Због тога се понекад примећују повећане концентрације летеће рибе у отвореним водама, чак стотинама миља низводно од акумулација фитопланктона на разилажењима.

Пре полетања у ваздух, „четверокрила“ летећа риба клизи дуж водене површине, убрзавајући своје кретање без вибрација репне пераје која остаје у води, а затим се одбија од површине и клизи летећи десетке и чак стотине метара.
Укупан број летећих риба у океанима је веома значајан. Према В.П.Шунтов, њихова залиха само у Тихом океану мери се у редоследу од 1,5-4 милиона тона, што је око 20-40 кг за сваки квадратни километар читавог тропског дела овог океана. Ове цифре су израчунате на основу резултата визуелног бројања риба које су излетеле испод стабла многих бродова у разним регионима и, по свему судећи, могу се приписати читавом Светском океану.
Број врста летећих риба у различитим регионима океана знатно варира, углавном због разлике у броју неритних и псеудо-океанских врста. Нарочито је много врста у водама Индонезије (27) и суседним регионима Коралног мора (26), у близини Филипинских острва (најмање 21) и јужног Јапана (25). Овде се налази - у западном тропском делу Тихог океана - модерно географско средиште ланца летећих риба и, по свему судећи, такође иницијално средиште формирања ове групе.
Поређење фауне летећих риба различитих делова Светског океана открива значајне разлике. Најбогатија и најразноврснија фауна летећих риба у Тихом океану, где постоји 47 врста и подврста. У Индијском океану до сада је пронађено само 26 врста, а у најтемељитије проученом Атлантском океану - само 16. Сваки океан има своје ендемске врсте, чији се број, међутим, знатно разликује. У Тихом океану живи 16 ендемских врста, у Индијском и Атлантику - свака по 4 ендемске врсте.
Треба напоменути да су све врсте ендемичне за Атлантик у Тихом и Индијском океану представљене врло сличним облицима. У исто време, многе групе врста су овде потпуно одсутне, уједињујући се у посебне подгенере и уобичајене у другим океанима. Генерално, фауна летећих риба у Атлантском океану је веома осиромашена (углавном због специјализованих врста подфамилије Ципселуринае, фауна индопацифичких летећих риба).
Летеће рибе Индијског океана и западног дела Тихог океана део су једне фаунистичке групе. Разлике у саставу врста летећих риба у различитим регионима тропског Индо-западног Пацифика објашњавају се углавном постојањем уско локализованих врста које заузимају ограничен домет. Ова фауна је најразноврснија и најкомплетнија у односу на родове и подгенере који су у њој заступљени (само род Фодиафор).
Фауна летећих риба у источном Пацифику врло је специфична. Обухвата не више од 20 врста, укључујући 9 ендемских врста и подврста. Ову фауну је са Атлантским океаном повезао род Фодиатор, али генерално изгледа да је сличнија индо-западно-пацифичком комплексу.
Тако се могу разликовати три главне географске групе летећих риба, које насељавају Индо-Пацифик, Источни Пацифик и Атлантски фаунистички регион. Може се претпоставити да су првобитни облици летећих риба настали у палеоцену или еоцену од предака блиских модерним полупужевима (породица Хемирхампхидае) у топловодним неристичким регионима који су историјски дуго постојали на граници Тихог и Индијског океани. Расејавање летећих риба из овог центра догодило се, очигледно, у свим правцима (али углавном према западу), иако његов пут још увек није довољно јасан.
Важну улогу у овом расејању очигледно је имао океан Тетис, кроз који су примитивни тропски елементи продрли у Атлантик заједно са осталим термофилним елементима. Екоцоетидае... Летеће рибе су несумњиво мигрирале кроз Панамски теснац, који је остао отворен до плиоцена, - само ово може објаснити савремену дистрибуцију рода Фодиафор на обе обале Централне Америке. Распрострањеност релативно „хладних“ суптропских риба догодила се, по свему судећи, много касније под климатским условима блиским савременим, а формирање биполарних подручја у њима, следећи теорију ЛС Берга, у потпуности се може објаснити променама у температурни режим океана током леденог доба.
Н.В.Парин
|









